Blue Brain Project
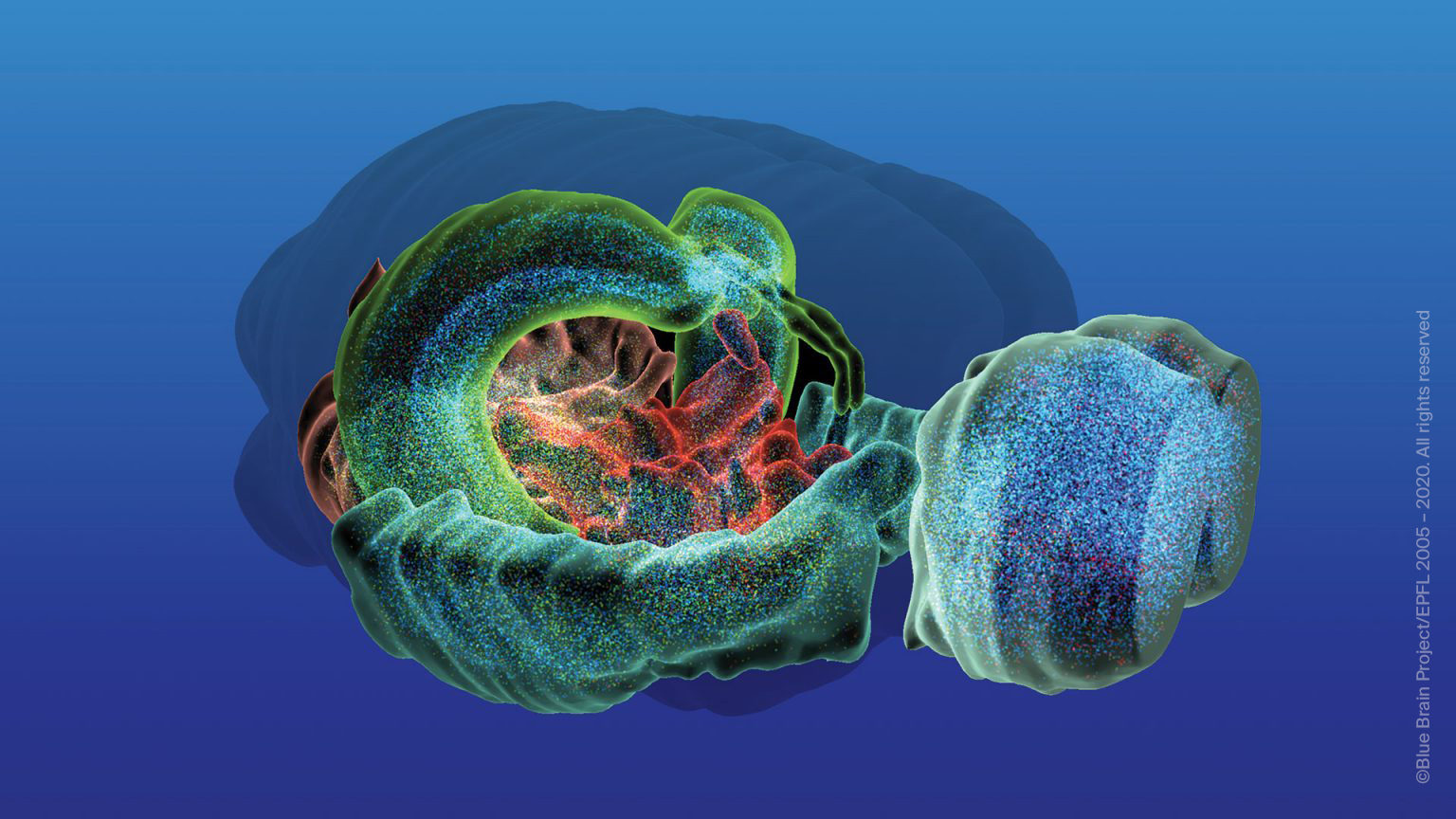
EPFL’s Blue Brain Project is a Swiss brain research Initiative led by Founder and Director Professor Henry Markram.
The aim of Blue Brain is to establish simulation neuroscience as a complementary approach alongside experimental, theoretical and clinical neuroscience to understanding the brain, by building the world’s first biologically detailed digital reconstructions and simulations of the mouse brain.
The supercomputer-based simulations and reconstructions built by Blue Brain offer a radically new approach for understanding the multi-level structure and function of the brain.


News
All the news about the Blue Brain Project
The Real Neuron Challenge

The Real Neuron Challenge
Can you tell the difference between a synthesized or biological neuron?

Events
See when the next Blue Brain Conference or Seminar will be held

Publications
Blue Brain has published 245 papers and pre-prints in scientific journals.

Blue Brain Portal
A knowledge space for simulation neuroscience


